| 平成10年度研究報告 VOL.48 香気性味噌酵母の開発 |
| |
3.結果と考察
3.1 TFL感受性試験
R株とD株のTFL濃度に対する感受性試験の結果をそれぞれ表2,表3に示す。R株では1〜2mMのTFL濃度で低頻度ながらコロニーが出現した。しかし,3mM以上のTFL濃度では完全に増殖が阻害された。また,D株については1〜3mMのTFL濃度でコロニーが出現し,4mM以上のTFL濃度では増殖が完全に阻害された。このようなコロニーの出現状況から,R株とD株はほぼ同等のTFL耐性を有するものと考えられた。この結果,いずれも低頻度ではあるが,自然誘発により1mM-TFLに耐性な株を取得できることがわかった。
表2 耐塩性酵母のTFL濃度に対する感受性
|
表3 耐塩性酵母のTFL濃度に対する感受性
| ||||||||||||||||||||||||||||||||
3.2 TFL耐性株の段階育種とイソアミルアルコール生成能
段階育種の結果,1〜4mMまでの各TFL濃度に耐性な株を段階的に取得することによって,最終的に5mMのTFL耐性株を取得することができた。ここで得られた5mM-TFL耐性株について,YPG-13培地による発酵試験を行い,イソアミルアルコール生成能を親株と比較した。その結果を図3に示す。
この結果,5mM-TFL耐性株のイソアミルアルコール生成能は親株と比較して,約1.5倍に増大し,香気生成能の向上が見られた。
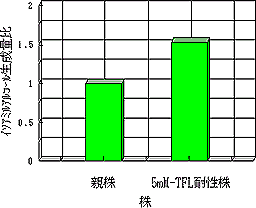 |
| 図3 TFL耐性株のイソアミルアルコール生成能 (親株の生成能を1とした) |
3.3 直接育種法によるイソアミルアルコール高生産株の育種
イソアミルアルコールを親株の2倍以上生成する株の取得を目的に,図4に示す操作法により,5mM-TFL培地にR株の培養液400μlを直接塗布して7〜10日間培養を行った。その結果,いくつかのコロニーが得られた(図5)。このときの出現頻度は,1.0〜3.6×10−7であった。同様に,D株についても1.6〜7.7×10−7の出現頻度で,5mM-TFL培地に生育可能なコロニーが得られた。この中から大きめのコロニー(3mm以上)を順に選抜し,新たに5mM-TFL培地へ画線培養を行った。この操作を5回繰り返し,最終的に生育の良好であった株を5mM-TFLに耐性な株として取得した。
この操作により得られたR株およびD株の5mM-TFL耐性株について,発酵試験による香気分析を行った結果,イソアミルアルコール生成能が親株の2倍以上を示す4株が得られた。これらをイソブタノールの生成量比とともに表4に示す。
これらの中で,R-5D-8株,D-5D-4株は,イソアミルアルコールだけでなく,イソブタノールについても親株の2倍以上を生成した。これらは,イソアミルアルコールだけを親株の2倍程度高く生成するR-5D-16株やD-5D-5株に比較して,官能的により強い芳香が感じられた。おそらくイソブタノールとイソアミルアルコールとの相乗効果により官能的に与える香りの強度が増大するものと考えられた。特に,D-5D-4株は,得られた株の中ではイソアミルアルコール,イソブタノールともに最高の生成量を示し,官能的にも良好な香気を示したので,D-5D-4株を香気性酵母として味噌の発酵試験(試醸)に用いた。
|
↓ 食塩5%YPD培地(5ml) 30℃,48hr,振とう培養 ↓ 培養液400μlを5mM-TFL培地へ直接塗布 ↓30℃,7〜10日間培養 コロニー ↓ コロニーを釣菌して5mM-TFL培地へ画線培養 ↓ ↓5回繰り返す ↓ ↓ 5mM-TFL耐性株(生育良好株) 図4 TFL耐性株の直接育種法 |
|
表4 TFL耐性酵母の香気生成試験
i-BuOH:イソブタノール 親株の各生成量を1とする。 |
3.4 味噌の発酵試験と試醸味噌の分析
香気性酵母D-5D-4添加区と対照の親酵母DY(P)添加区及び酵母無添加区の3種類の仕込み区で,90日間熟成した各味噌の水分,食塩,pHの分析結果を,仕込み直後の値(括弧内の数値)とともに表5に示す。いずれも仕込み直後に比較して,若干の水分減少と塩分濃度の増加が認められたが,ほぼ同程度の値であった。
また,各味噌のその他の一般成分を表6に示す。糖分値では,酵母添加区ではいずれも酵母無添加区に比較して低い値を示した。これは,酵母の糖分消費によるものと考えられた。その他の成分については,各味噌の間で大きな差異は見られなかった。
表5 味噌の分析結果
|
測定項目
区 |
水分 (%) |
食塩 (%) |
pH |
| 酵母無添加区 DY(P) 添加区 D-5D-4 添加区 |
41.1(45.5) 42.3(45.4) 42.0(45.6) |
13.3(12.6) 13.3(12.7) 13.3(12.8) |
5.00(5.98) 4.95(5.98) 5.03(6.00) |
熟成日数:90日,( )内は仕込み直後の分析値
3.5 酵母生菌数
熟成期間中の各味噌の酵母生菌数変化を表7に示す。香気性酵母D-5D-4及び親酵母DY(P)添加区では,いずれも25日目に4.3×106/gの最高菌数に達し,良好な増殖を示した。また,酵母無添加区では,18日目まで102/g未満であったが,25日目以降,次第に増加し,90日目には106/gオーダーまで増加し,その後次第に減少した。3.8で述べるように,酵母無添加味噌のエタノール濃度が0.1%程度と低く,温度を15℃に低下させた54日目ごろから増殖が盛んになり,高濃度食塩(18%NaCl)を添加した培地中のマルトースを資化した26)ことなどから,この酵母は後熟酵母のCandida属と推定される。
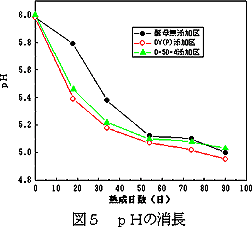
3.6 熟成期間中の味噌のpH変化
熟成期間中の各味噌のpH変化を図6に示す。DY(P)及びD-5D-4酵母添加区では,酵母無添加区に比較して初期のpHの低下が大きかった。これは,発酵に伴う炭酸ガスの発生によるところが大きい27)と考えられる。その後,90日目ごろに,いずれの仕込み区の味噌もほぼpH5付近に達し,熟成の終了時期と判断した。
表6 味噌の成分分析結果
|
測定項目
区 |
全糖 (%) |
直糖 (%) |
全窒素 (%) |
水溶性 窒素(%) |
フォルモール 窒素(%) |
酸度Ⅰ (ml) |
酸度Ⅱ (ml) |
| 酵母無添加区 DY(P) 添加区 D-5D-4 添加区 |
21.9 18.1 18.3 |
18.4 15.4 15.4 |
1.68 1.68 1.68 |
0.96 0.99 0.99 |
0.35 0.36 0.36 |
11.6 11.7 11.3 |
11.4 11.1 10.8 |
熟成日数:90日
表7 味噌中の酵母生菌数変化(cells/g)
|
熟成日数
区 |
0 | 18 | 25 | 34 | 54 | 74 | 90 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 酵母無添加区 DY(P) 添加区 D-5D-4 添加区 |
|
|
|
|
|
|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 |
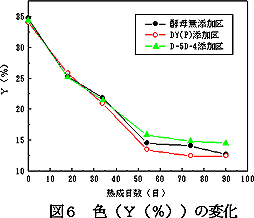 |
| 図6 熟成期間中の味噌のpH変化 | 図7 味噌の色(Y(%))の変化 |
3.7 熟成期間中の味噌の色変化
熟成期間中の味噌の表面色(Y(%))の変化を図7に示す。35日目ごろまでは,いずれも大きな差はなかった。しかし,それ以降,DY(P)酵母添加区では,色がやや早く濃化した。90日目の香気性酵母D-5D-4添加の味噌のY(%)は14.5%で,酵母無添加の12.7%,親酵母DY(P)添加の12.4%よりも2%程度高く,比較的淡色傾向にあった。
3.8 香気成分
90日目の各味噌のエタノールは,香気性酵母及び親酵母の添加区では,いずれも0.8%程度,また,酵母無添加区では,90日目でも0.1%程度であった。
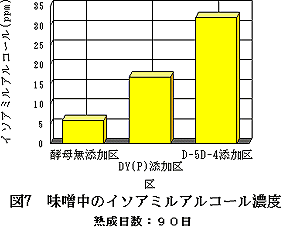
味噌の香気成分のうち,90日目の各味噌の生成イソアミルアルコール濃度を図8に示す。D-5D-4酵母の添加区では約32ppmを示し,親酵母DY(P)添加区の約17ppm,酵母無添加区の約6ppmと比較すると,それぞれ約1.9倍,約5.3倍高い値であった。この結果,D-5D-4は味噌の発酵においても,親酵母の約2倍のイソアミルアルコール生成能を反映した。
また,90日目の各味噌の生成n-ブタノール濃度を図9に示す。酵母無添加区では,n-ブタノールの生成がほとんど認められなかったのに対し,D-5D-4添加区では10.5ppmを示し,DY(P)添加区の5.7ppmと比較すると,約1.8倍高いn-ブタノール生成能を示した。
n-ブタノールは,液体培養では生成されず,味噌のような半嫌気的な状態でのみ生成される特異な成分である。大西ら28)は全高級アルコールに占めるn-ブタノールの比率が高い味噌ほど,香りの評価が高いとしている。したがって,ここで得られた香気性酵母添加の味噌は,香りに対する評価の高いことが期待される。一方,他の香気成分n-プロパノール,イソブタノールについては,D-5D-4酵母添加の味噌は,親酵母DY(P)添加の味噌と比較して大きな差は認められなかった。以上の結果より,香気性酵母D-5D-4を添加した味噌は,親酵母DY(P)を添加した味噌と比較して,イソアミルアルコールでは約1.9倍,n-ブタノールでは約1.8倍高い値を示し,味噌の香気成分,高級アルコールの増大に効果が認められた。
酵母がTFL耐性を獲得することによりイソアミルアルコール生成能が増大することについては,最近,S.cerevisiaeのTFL耐性株に対して遺伝子的解析がなされ29),30),TFL耐性酵母はグルコースからロイシンの生合成系の律速酵素であるα-イソプロピルリンゴ酸(α-IPM)シンターゼが変異を受けていることが明らかとなった。すなわち,従来考えられていたように,ロイシンのフィードバック阻害に非感受性で,結果的にイソアミルアルコール生成の酵素活性が増大したものと考えられた。さらに,変異α-IPMシンターゼは,Zn2+介在型CoAによる失活に対しても耐性であり,ロイシンの代謝系がCoAレベルで制御されていることをも示唆した。おそらく,同様のことがZ.rouxiiに対しても考えられるのではないかと推察される。
 |
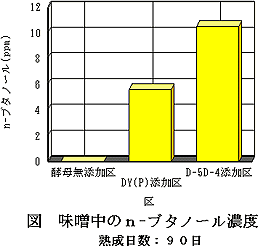 |
| 図8 味噌中のイソアミルアルコール濃度熟成日数:90日 | 図9 味噌中のn−ブタノール濃度熟成日数:90日 |
3.9 官能審査
味噌の官能審査について,t−検定法を用いて統計処理した結果を表8に示す。香気性酵母添加の味噌は,色,味,香り,総合評価のすべてにおいて酵母無添加の味噌との間に有意な差が認められた。
表8 官能審査(t検定)
| 項目 | 区 |
|
|
|
|
| ||||||||||
| 色 | 酵母無添加区(A) DY(P)添加区(B) D-5D-4添加区(C) |
1.85 2.70 1.45 |
0.39 0.42 0.41 |
|
|
| ||||||||||
| 香り | (A) (B) (C) |
2.30 1.98 1.73 |
0.68 0.59 0.61 |
1.83 |
|
1.44 | ||||||||||
| 味 | (A) (B) (C) |
2.38 2.05 1.58 |
0.50 0.77 0.46 |
1.83 |
|
| ||||||||||
| 総合 評価 |
(A) (B) (C) |
2.38 2.18 1.45 |
0.45 0.66 0.46 |
1.20 |
|
|
パネラー:40名、**:危険率1%で有意
味噌の官能審査のうち,総合評価について行った結果を図10に示す。色,味,香りの総合評価で,40名中26名が香気性酵母添加の味噌に最良点を与え,親酵母添加の味噌の10名,酵母無添加の味噌の4名を大きく上回った。この結果,本研究で得られた香気性酵母添加の味噌が高く評価されたことから,全香気成分に占める高級アルコール分の多い味噌ほど評価が高いということが官能的にも裏付けられた。また,味噌中の香気成分も増大したことから,味噌への応用が十分可能であることがわかった。
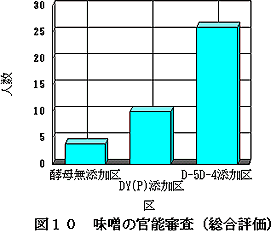 |
| 図10 味噌の官能審査(総合評価) |
4.結言
味噌・醤油用耐塩性酵母から,アミノ酸アナログ5,5,5-トリフルオロロイシン(TFL)に耐性な株を育種し,味噌の香気成分の一つ,イソアミルアルコール生成能を高めた株を取得した。TFL耐性酵母の育種は,1mM-TFLを含む最小培地に,自然誘発により生育してきたコロニーをもとに,TFL濃度を順次高め,最終的に5mM濃度の培地まで段階的に酵母を植え継いでいく段階育種法と,酵母の培養液を直接5mM-TFL含有培地に塗布する直接育種法で行った。その結果,5mM-TFL耐性株の中から,段階育種法では親株に比較して約1.5倍量の,さらに,直接育種法では,取得株の中では最高の約2.6倍量のイソアミルアルコール生成能を示す株が得られた。そこで,後者の香気性酵母を用いて,5kgスケール,90日間の味噌の試醸を,酵母無添加,親酵母添加の場合と比較して行った。その結果,(1)香気性酵母を添加した味噌のイソアミルアルコール含有量は,酵母無添加味噌の約5.4倍,親酵母添加味噌の約1.9倍高い値を示した。(2)n-ブタノールについては,香気性酵母添加味噌では親酵母添加味噌の約1.8倍高い値を示した。この結果,香気性酵母の添加により高級アルコールが増大し,味噌の香気向上に効果が認められ
た。(3)官能審査では,色,味,香りを含めた総合評価において,40名中26名が香気性酵母添加の味噌に最良点を与え,親酵母添加の10名,酵母無添加の4名を大きく上回った。香気性酵母添加の味噌が高く評価されたことは,全香気成分に占める高級アルコールの量が多い味噌ほど評価が高いことを官能的にも支持し,味噌への応用が十分可能であることを示唆した。
謝辞
本報をまとめるにあたり,適切なご助言を頂いた金沢大学理学部福森義宏教授に感謝します。
参考文献
1)安平仁美:醸協,Vol.75, No.6, p.506-515(1980).
2)高橋清:醸協,Vol.92, No.4, p.241-250(1997).
3)渡辺隆幸:醸協,Vol.93, No.1, p.22-27(1998).
4)吉川茂利,小栗勇:醸協,Vol.87, No.11, p.780-787(1992).
5)末沢保彦,池田知子:香川県発酵食品試験場研究報告,No.84, p.23-27(1991).
6)S. Ashida, E. Ichikawa, K. Suginami and S. Imayasu: Agric. Biol. Chem.,
Vol.51, No.8,p.2061-2065(1987).
7)市川英治:醸協,Vol.84, No.3, p.166-170(1989).
8)秋田修,蓮尾徹夫,原昌道,吉沢淑:発酵工学,Vol.67, No.1, p.7-14(1989).
9)K. Fukuda, M. Watanabe, K. Asano, H. Ueda and S. Ohta : Agric. Biol.
Chem., Vol.54, No.1, p.269-271(1990).
10)秋田修,井田哲朗,小幡孝之,原昌道:醸協,Vol.85, No.7, p.501-505(1990).
11)O. Akita, T. Ida, T. Obata and S. Hara : J.Ferment. Bioeng., Vol.69,
No.2, p.125-128(1990).
12)青木光達,内田金治:農化, Vol.62, No.3, p.481(1988).
13)中村誠:日食工誌,Vol.37, No.12, p.981-983(1990).
14)T. Aoki and K. Uchida : Agric. Biol. Chem., Vol.55, No.11, p.2893-2894(1991).
15)伊藤公雄,武田茂,中村昌昭,早出昭雄,今井学,安平仁美:信州味噌研究所研究報告,
No.34, p.7-13 (1993).
16)S. Yoshikawa, I. Oguri, K. Kondo, M. Fukuzawa, M. Shimosaka, M. Okazaki:
FEMS Microbiol. Lett., Vol.127, p.139-143(1995).
17)久寿米木一裕,出永直樹,吉田秀恵:醸協,Vol.92, No.11, p.827-834(1997).
18)久寿米木一裕,出永直樹,吉田秀恵:醸協,Vol.92, No.11, p.842-845(1997).
19)久寿米木一裕,馬場雅幸,吉田秀恵,宮城盛安:生物工学, Vol.75, No.5, p.343-346(1997).
20)久寿米木一裕:醸協,Vol.93, No.8, p.606-614(1998).
21)近藤君夫,吉川茂利,蟻川幸彦,桑原秀明,小栗勇:長野食工試研報, No.17, p.43-47(1989).
22)吉川茂利,近藤君夫,桑原秀明,蟻川幸彦,小栗勇,岡崎光雄:長野食工試研報,
Nol.18, p.71-76(1990).
23)全国味噌技術会編:味噌技術ハンドブック(1995).
24)全国味噌技術会編:基準味噌分析法(1968).
25)全国味噌技術会編:みそ製造技能士のハンドブック(1979).
26)三輪昭生,伊藤寛,姫野国夫,新国佐幸:醤研, Vol.15, No.3, p.86-92(1989).
27)松本伊左尾,今井誠一:醸協,Vol.69, No.9,p.590-594(1974).
28)大西邦夫,安平仁美:昭和58年度日本発酵工学会講演要旨集,p.33-34(1983).
29)E. Casalone, G. Fia, C. Barberio, D. Cavalieri,L. Turbanti and M. Polsinelli:
Res. Microbiol.,Vol.148, p.613-623(1997).
30)D. Cavalieri, E. Casalone, B. Bendoni, G. Fia,M. Polsinelli and C. Barberio:
Mol. Gen. Genet.,Vol.261, p.152-160(1999).
|
|
|